2020. aastal võitis Soome maineka Finlandia kirjandusauhinna Anni Kytömäki romaani „Margarita“ eest [6]. Bioloogi haridusega autor oli mitme aasta jooksul tegelenud ebapärlikarbiga ja raamatu pealkiri tulenebki ebapärlikarbi (Margaritifera margaritifera) varasemast ladinakeelsest nimetusest Margarita. Ebapärlikarp kannab selles teoses ka üht olulist teemaliini.
Tekst ja fotod: KATRIN KALDMA

Mis teeb mõne liigi inimese meelest erilisemaks ja huviäratavamaks kui teise? Harilikult köidavad liigi iseärasused, mida saab võrrelda inimese omadega. Ebapärlikarbil on näiteks pikk eluiga ja väidetavalt niisama kaua kestev paljunemisvõime. Ent tõeliselt paeluvaks teeb selle liigi pealtnäha lihtsa välimuse taga peituv keeruline eluring ja veekeskkonnast tublisti kaugemale ulatuv „suhtevõrgustik“.
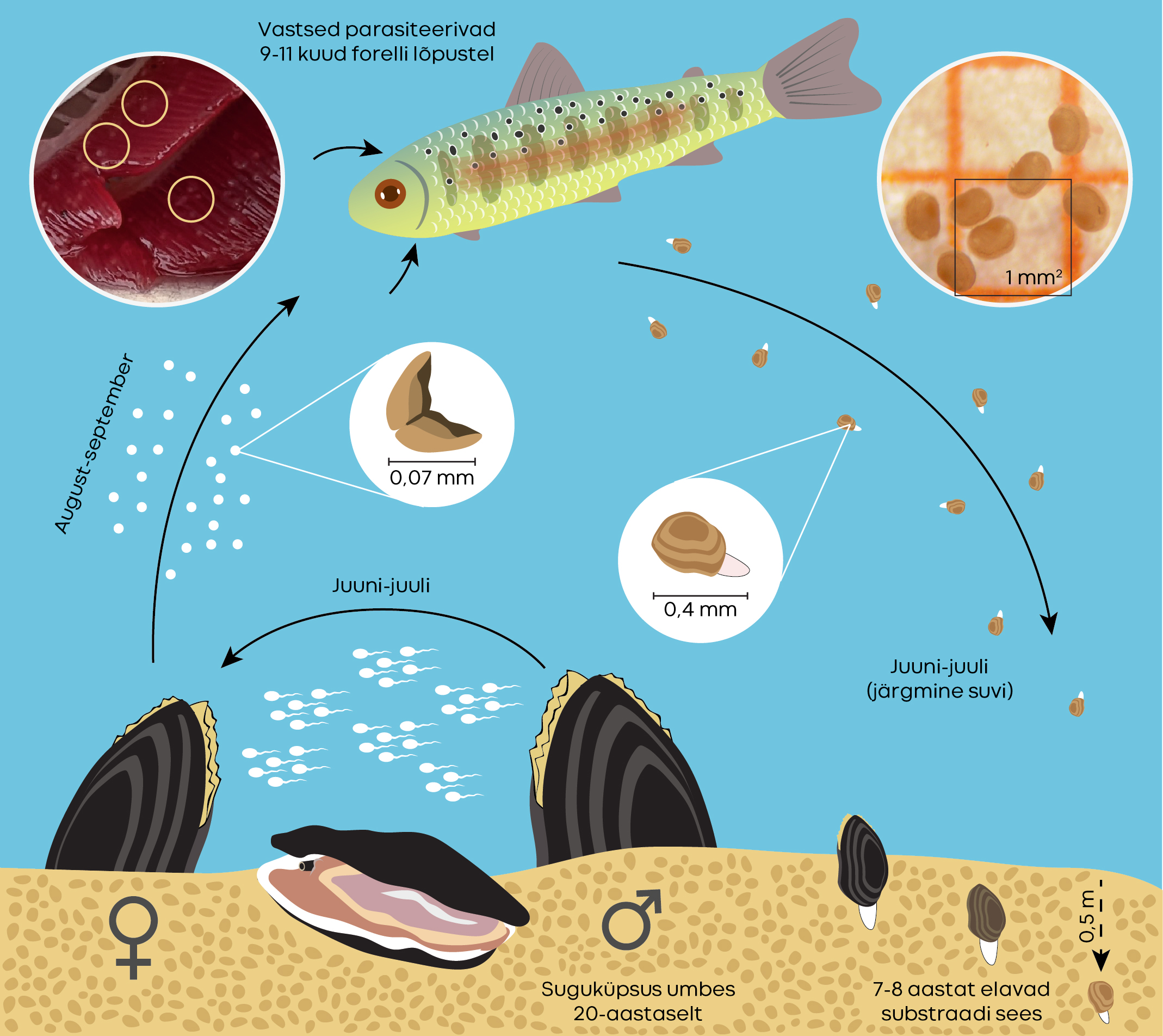
Ebapärlikarbi täiskasvanud isendid asetsevad jõepõhjas liikumatult reas nagu kivid. Nad ei reeda millegagi, et nende väheliikuva ja pika elu algust iseloomustab hulk loteriilaadseid seiku, millest oleneb järelkasvu saatus.
Elukäik õnnestub, kui
• isasloomadest vette väljutatud seemnerakud jõuavad viljastumisjärgus emasloomani,
• vastsed paiskuvad emasloomast välja sel ajal, kui lähikonnas ujuvad noored kalad,
• vastsed jõuavad kinnituda sobiva peremeeskala lõpustele või pudenevad jõepõhja,
• nakatunud, st vastseid kandvad kalad pääsevad eluga kõikide kalatoiduliste käest, hoolimata kala noorjärkude suurest suremusest,
• kaladelt vabanenud noored karbid kukuvad sobivale jõepõhjale, kuhu on võimalik kaevuda (kuni 30 cm sügavusele),
• vähemalt viis kuni kaheksa järgmist aastat on jõepõhi selles lõigus settest puhas ja vesi hapnikurikas.
Kalast peremees
Karbivastsed reisivad esimestel elukuudel parasiitse järguna kala lõpustel ja sõltuvad täielikult oma peremeesorganismist, st nad on obligatoorsed parasiidid. Ebapärlikarbi peremeesorganism on lõhe või forell, Pudisoo jões on peremees forell. Kõige paremini sobivad forellid nende esimestel eluaastatel, eelkõige samal kevadel koorunud isendid, sest vanemaks saades kalade nakatumisvõime kahaneb [10]. Katseoludes suudavad vastsed kinnituda nii forellile kui ka lõhele, kuid looduslikes tingimustes leidub nendes jõgedes, kus elavad mõlemat liiki kalad, ka kahte laadi karpe: ühed parasiteerivad forellil ja teised lõhel. Vastsete küpsemise aeg lõhe- või forelliparasiitide vanematel erineb
paar nädalat; forellil parasiteeriva nn osapopulatsiooni emakarbid paiskavad vastsed välja varem (kuni neli nädalat)
kui lõhel parasiteeriva osapopulatsiooni emased [5].
Kuidas parasiteerimine peremeest mõjutab? Seda on üksjagu uuritud. Väike kogus parasiite kalade käitumist ei muuda, aga raskelt nakatunud kalad on siiski halvemas seisundis kui nende nakatumata kaaslased ja hukkuvad sagedamini röövluse või muude asjaolude tõttu [2]. Karbivastseid kandvate lõhilaste reaktsiooniaeg on pikem ja liikumiskiirus väiksem kui tervete samavanuste liigikaaslaste oma. Esimese talve veedavad karbivastsed lõpustel. Mida pikemaks kalast sõltuv ajajärk kujuneb, seda parema ettevalmistuse iseseisvaks karbieluks vastsed saavad [8]. Ebapärlikarbi levila piirides on see aeg küllaltki erinev ja oleneb kõikide päevade summaarsest veetemperatuurist: kui ilmad on külmad, siis kulub valmimiseks rohkem aega, kui ilmad (ja vastavalt vesi) on soojemad, nt talv pehmem, kevad varasem, siis valmivad vastsed kalalõpustel varem ja pudenevad kiiremini [12].
Meie karbid kinnituvad pihtvastsena kalale (lõpustele) augustis või septembris ja lahkuvad kalast karbikujuliste noorloomadena järgmisel suvel. Selle ligikaudu kümne kuu jooksul kulgeb kasv kahes etapis.
Esimene järk kestab talviste külmade tulekuni. Teine etapp algab, kui kevadine vesi on soojenenud, ja kestab, kuni saavutatakse õige kuju ja suurus (umbes 0,4 mm). See ei tähenda igavat vaikelu. Tähelepanelikud uurijad on teinud kindlaks kolm staadiumi, mille vältel parasiitsed karbihakatised võtavad järgemööda pähkli, ümara oa ja tavapärase oa kuju [1].
Kui kõik läheb hästi ja karbihakatised satuvad kaevumiseks sobivasse jõelõiku, siis jätkub nende loomulik areng ja kasv jõepõhjas. Paraku hääbuvad paljud Euroopa asurkonnad just selle pika ahela viimase lüli nõrkuse pärast. Jõgedes napib karbivastsetele sobivat jämeliivast ja peenekruusast jõepõhja, kus hapnikurikas vesi voolaks vabalt läbi põhjakihi ja toidaks sealseid karpide lasteaedu.
Elu jõepõhjas
Jõepõhi iseseisva elu esimese kasvukeskkonnana katab mitmeid vajadusi. Kuigi kaladelt pudenevad jõkke justkui täiskasvanute tillukesed koopiad, ei ole need kõige „karbilikuma“ omaduse poolest veel täisväärtuslikud. Verinoored karbid ei olegi filtreerijad, vaid toituvad jala abil [7]. Üleminekuiga algab, kui on ületatud ligikaudu ühemillimeetrine mõõdulatt. Elu jõepõhja kivide vahel võimaldab hakkama saada kiire voolu ja tulvadega, kividele tekkivat biokilet ja kogunenud pudemeid kasutavad noored karbid toiduks. Ühtlasi kaitseb sealne mikrokliima väikseid organisme talvel külmumise eest.
Ebapärlikarbi vastse- ja „noorukiea“ kestust saab võrreldav inimese asjaomaste eluetappidega. Pärlikarbid kasvavad väga aeglaselt: loodusoludes ligikaudu millimeetri aastas. Kui võrreldi eri Hispaania jõgedes elavate karpide kasvu kiirust, ilmnes, et esimeste aastate veidi kiirem kasv aeglustub pärast kuuendat eluaastat [11]. Siin kandis ja mujal levila põhjaosas kasvavad pärlikarbid pigem veel aeglasemalt ja enamasti jääb kasvulaeks 11 või 12 sentimeetrit. Samas ei teata lage isendite vanusel. Pärlikarbimaailmas ei ole midagi erilist elada üle 150 aasta vanaks, endistviisi tuleb kinnitatud või kinnitamata teateid uute vanuserekordite kohta. Soome teadaolevalt vanim karp oli üle 200 aasta vana.
Karpide kasvu mõjutab toitainete kogus jões. Ühepikkuste poolmete vahel võivad elada nii 70 kui 120 aasta vanused isendid. Kahjuks osatakse nende vanust täpselt määrata ainult ühe meetodi järgi: värvitakse karbikoja läbilõige ja sealt loetakse aastaringide hulk. See aga tähendab, et iga kord, kui on hinnatud uuritava isend vanust, on tema pikk elu inimese uudishimu tõttu saanud lõpu.

Kui palju kulub aega, enne kui siinsed noored karbid hakkavad järglasi andma, pole täpselt teada. Inglismaal on see 12 aastat, samas Tšehhis kulus 20 aastat, enne kui ajakohaste tugisüsteemide abil üles kasvatatud ja loodusesse tagasi viidud pärlikarp hakkas sigima (Ondřej Spisar, suuline teade). Rahvusvaheline looduskaitseliit (IUCN) on selle liigi ühe põlvkonna pikkuseks arvestanud 30 aastat [10].
Iidsete aegade asukas
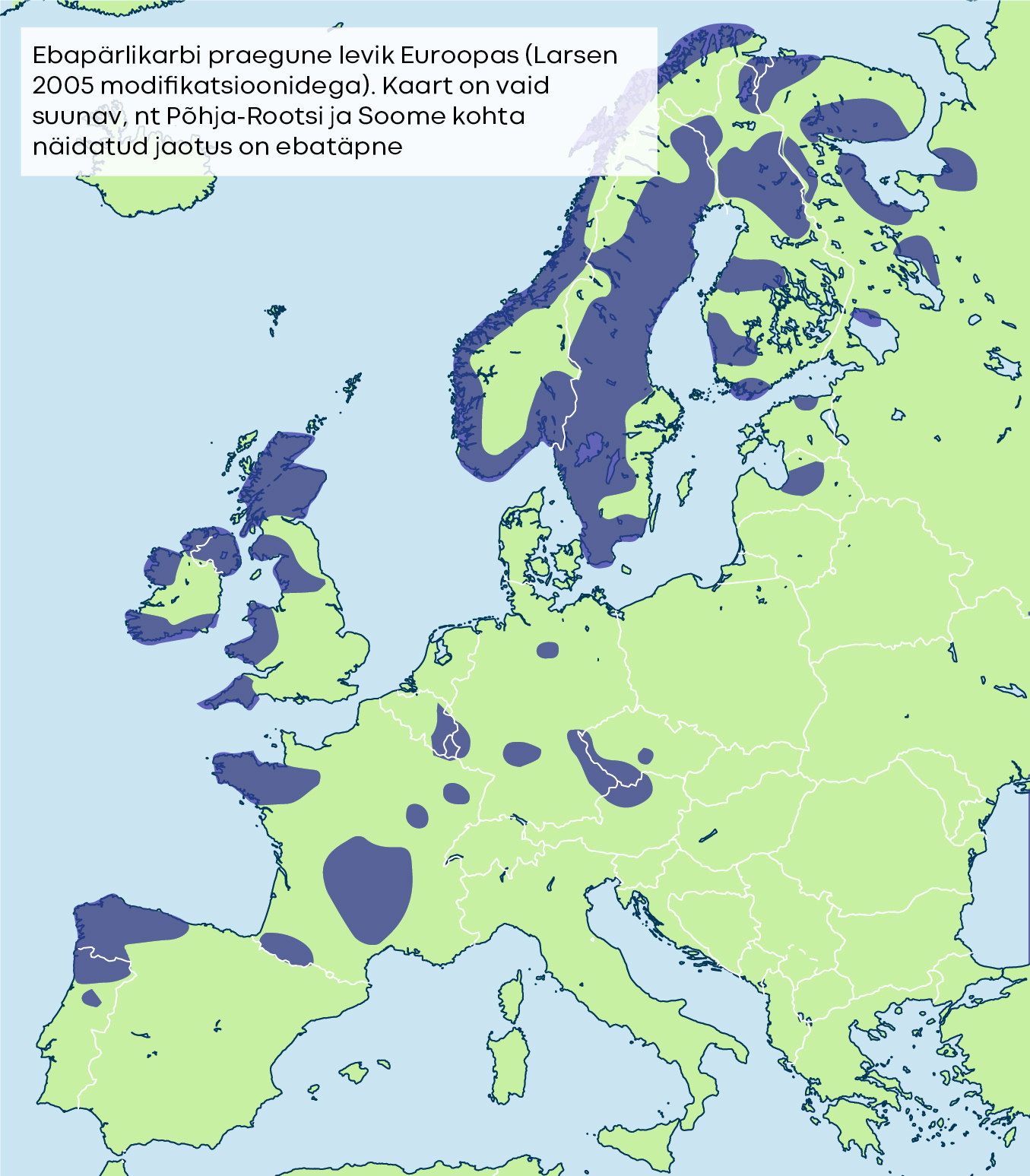
Kõnealune liik hakkas kujunema Kriidi ajastul 100 miljonit aastat tagasi. Nüüdsel ajal on ta levinud põhjapoolkera jahedates toitainevaestes ja kiirevoolulistes jõgedes. Lõunapoolsemad karbijõed asuvad Hispaanias ja Portugalis, põhjapoolsemad Põhja-Soomes ja Norras. Liigile kõige olulisemad alad sadade elupaigajõgedega paiknevad Norras, Soomes ja Suurbritannias.
Eestis ei olegi peale praeguse elupaiga olnud kuigi palju karbijõgesid. Kolme sajandi taha ulatuvate teadete järgi, mille eest võlgneme tänu pärlipüüdjatele, oli tolleaegsel Liivimaal, praegusel Lõuna-Eesti alal, veel paar pärlikarbijõge ja Põhja-Eestis veel üks järv, kus seda liiki leidus [13].
Paljude teiste Põhja-Eesti jõgede suur kaltsiumisisaldus välistab need veekogud võimalike ebapärlikarbile sobivate elupaikade nimistust. Veetemperatuur, pH, kaltsiumisisaldus ja lõhilased ongi need põhilised parameetrid, mis määravad jõe sobivuse liigi elupaigana. Laia levila piirides on iga pärlikarbijõgi ja selle asurkond ainulaadne pikalt kokku mänginud koosseis.
Jõgesid ja ojasid iseloomustab voolurežiim, milles pole kuigivõrd korrapära, kuid on siiski selged kordused, mille mõjul on kujunenud elu vees ja kallastel. Kauaaegse kooseksisteerimise tulemusel on moodustunud pikad suhteahelad ja võrgustikud. Praeguste teadmiste põhjal ei vali täiskasvanud ebapärlikarbid toitu. Filtreerivatele ja väheliikuvatele loomadele satub suhu see, mida kannab 50 liitrit jõevett, mis täiskasvanud isendit ööpäevas läbib. Kõik, mida on võimalik omastada, kasutatakse ära elutegevuseks, st väga aeglaseks kasvuks.
Ülejäägid väljutatakse ja leiavad tee mõne teise organismi toidulauale ja talletatakse selle kasvamises. Mitmesugused vette sattuvad või seal tekkivad ühendid, mis kalad või teised veeorganismid ei suuda esialgu omastada, muutuvad karbi vahendusel neile kättesaadavaks.
Ei saa hakkama detriidita
Toitaineid valgub vette nii lähedastelt märgaladelt kui ka jõkke voolavatest ojadest ja allikatest. Ka sademete tagajärjel tekkivad ajutised sissevoolud toovad toitaineid. Oluline komponent on osakesteks lagunenud surnud organismide jäänused: pude ehk detriit. Populatsiooni taastamise keskustes on tehtud kindlaks, et noorkarpide elu ilma detriidita ei ole võimalik.
Varasemate uuringute järgi peeti kõige tähtsamaks jõeäärsete alade, st väljastpoolt jõge sisse kandunud (allohtoonset) detriiti [4]. Värskema uuringu põhjal selgus, et rohkem väärtuslikke polüküllastunud rasvhappeid sisaldas jões kohapeal tekkinud (autohtoonne) detriit, mis tagas kiirema kasvu ja parema ellujäämuse [3].
Siiski oletatakse, et mõlemad komponendid etendavad suurt rolli. Väljastpoolt sisse kandunud detriidi väärindavad jões olevad ränivetikad, kottseened ja teised organismid jõedetriidiks, mis on eelkõige ebapärlikarbi noorjärkudele toiduks äärmiselt vajalik. Detriidi teine suur roll on olla jõkke ajuti kogunevate toksiliste ühendite puhverdaja. Seetõttu on pärlikarbi ja teiste jõeelanike eluks ülitähtis, et nii kaldaalade kui ka jõe enda laguahelad toimiksid.

Mis ohustab ebapärlikarpi?
Loodus teeb kõik, et ebapärlikarpe oleks jõgedes palju. Igal aastal paiskavad täiskasvanud karbid välja miljoneid vastseid. Ent sellest ei piisa, kui elupaigaga pole kõik korras.
Ebapärlikarp on selgeveelise jõe liik. Puhas vesi ja uued veevarud saavad tekkida aeglase filtreerimisprotsessi tulemusel. See tähendab, et vesi peab jõudma maastikule ja jääma sinna võimalikult kauaks. Üks peamine vooluveekogude taastamise võte ongi aeglustada vee liikumist.
Võrreldes meie teiste jõgedega leidub ebapärlikarbijõel ilusaid pikki looduslikke lõike, kus voolusängi pole rikutud. Kuid jõe ülemjooksul, lähikonna rabades ja märgades metsades on rikutud veerežiimi. Jõkke suubuvad ojad on kaevatud sirgeteks kuivenduskraavideks. Igasugune kraavide rajamine või värskendamine muudab kohe vee koostist ja vee-elustiku toidulauda.
Kuivendussüsteemide kaudu sängi lisandunud setted ummistavad noorte karpide ellujäämiseks vajalikku puhast põhjakruusa ja rikuvad elupaiga mitmekesisust. Kui valgala loomulikku veerežiimi on muudetud, halveneb tunduvalt jõe isepuhastumisvõime ja väheneb üldiselt veekogu panus põhjaveereservi taastumisse. Kuivendamise tõttu teisenevad kaldakooslused, sh muutuvad laguahelad.
Jõge ületavad teed ja iga truup, mis jõkke asetatakse, lõhub loodusliku elupaiga terviklikkuse, halvimal juhul takistab veeliikide liikumist. Otseste reostusallikatega tegeletakse aasta-aastalt üha põhjalikumalt, kuid täielikult tähelepanuta on jäänud talvise teehoolduse mõju looduslikele veekogudele. Seda võimendavad tuhanded kilomeetrid maanteekraave. Endiselt ohustab ebapärlikarbi püsimajäämist peremeeskalade vähesus, eelkõige forelli röövpüügi tõttu.
Teiste liikide mõju
Mujal Euroopas on ebapärlikarbi võimaliku loodusliku ohutegurina nimetatud signaalvähki, kes piisava asustustiheduse korral suudab karbikodasid vigastada [11]. Leidub üksjagu loomi, kes toituvad väikesemõõdulistest kaladest. Kui peremeeskalade arvukus jões millegi tõttu kahaneb, siis võib tekkida konflikt eri kaitsealuste liikide vahel. Mitmes Euroopa riigis on nii juhtunud saarma ja ebapärlikarbiga.
Kopra tegevus loodusmaastikus on mitmeti positiivne ja ebapärlikarbile ta seal ohtu ei kujuta. Paisutus mõjub sel juhul nagu suurvesi, mis tõuseb kallastele ja jätab lahkudes palju jõesetteid maha. Kobras loob märgalasid ja teisi uusi elupaiku, mis sobivad paljudele liikidele. Tema tegevus mitmekesistab maastikku kõikvõimalikul moel.
Ent kui veerežiim on rikutud ja jõgi tasakaaluseisundist välja viidud, mõjutab kobras habrast tasakaalu veelgi enam. Kaldaerosiooni tõttu kaob peavoolusäng, juurealused kui olulised elupaigaosad täituvad settega, moodustub kausjas mudane jõesäng peaaegu seisuveeliste lõikudega. Vähene vesi hajub ühetaolisel jõepõhjal ja täiskasvanud karbid jäävad kuivale.
Suvel hakkab säärastel lagedatel veealadel vesi soojenema ja soodustab vetikate kasvu. Veetemperatuuri tõus ja hapnikusisalduse vähenemine on kriitiline nii forellile kui ka karpidele ja avaldab mõju mitukümmend meetrit allavoolu. Juba mõnekraadine vee soojenemine sobib hästi kalaparasiitidele jt haigustekitajatele. Jõelähedaste poollooduslike koosluste korral raskendab kobras oma elutegevusega märkimisväärselt nende koosluste hooldust või isegi välistab selle.
Karpide käekäik kajastab ürglooduse seisundit
Millises jões eelistab ebapärlikarp elada? Arvatakse, et parimad lõigud paiknevad allikaliste alade läheduses. Allikad hoiavad ühtlasemat veetemperatuuri. Temperatuuri mõju on arvatavasti kõige kriitilisem varases arengujärgus, kui noorte ebapärlikarpide energiavarud on talve hakul väikesed. Kui jõevesi jahtub nullilähedale, siis allikatest sisse voolava vee temperatuur jääb talvel mitu kraadi kõrgemaks. Ja nagu eespool öeldud, toovad allikad jõkke detriiti. Niisugustes parimates jõelõikudes võivad pärlikarbid ise ollagi jõepõhi, paiknedes kiht-kihilt üksteise peal, kohati kuni mitmesaja kaupa ruutmeetril.
Mida peame pidama silmas, kui räägime ebapärlikarbist? Jahedaid selgeveelisi jõgesid ja ojasid. Ürgseid loodusmaastikke, kus need jõed-ojad voolavad. Põnevaid ja suurel määral tundmatuid võrgustikke, mis ulatuvad äärmiselt kaugele.
Elu, ka ebapärlikarbi oma, on võrgustik, mille toimimine eeldab tervet ja terviklikku ökosüsteemi. Kui karpide seisund on halvenenud või on need loodusest kadunud, võime olla kindlad, et meile kõigile olulise looduskeskkonna seisund on muutunud halvemaks.
Sada miljonit aastat pikk eksistents peaks olema piisav tõestus selle kohta, et liik on kohane elama ajas muutuvas keskkonnas. Ometi oleme viimasel sajandil teinud midagi nii valesti, et väga pikka aega Maal elanud loomast on saanud üks Euroopa ohustatuim mageveeliik.
- Castrillo, Pedro A. et al. 2022. The parasitic travel of Margaritifera margaritifera in Atlantic salmon gills: from glochidium to post-larva. – Aquaculture Reports 27:101340. https://doi.org/10.1016/j.aqrep.2022.101340.
- Filipsson, Karl et al. 2018. Heavy loads of parasitic freshwater pearl mussel (Margaritifera margaritifera L.) larvae impair foraging, activity and dominance performance in juvenile brown trout (Salmo trutta L.). – Ecology of Freshwater Fish. 27. 70-77. 10.1111/eff.12324.
- Grunicke, Felix et al. 2023. Riparian detritus vs. stream detritus: food quality determines fitness of juveniles of the highly endangered freshwater pearl mussels.
- Hruška, Jaroslav 1995. Problematik der Rettung ausgewahlter oligotropher Gewassersysteme und deren natürlicher Lebensgemeinschaften in der Tschechischen Republik. – Lindberger Hefte 5: 98–123
- Karlsson, Sten et al. 2014. Host-dependent genetic variation in freshwater pearl mussel (Margaritifera margaritifera L.). – Hydrobiologia 735: 179–190. doi:10.1007/s10750-013-1679-2.
- Kytömäki, Anni 2021. Margarita. Ühinenud Ajakirjad, Vesta.
- Lavictoire, Louise et al. 2018. Ontogeny of juvenile freshwater pearl mussels, Margaritifera margaritifera (Bivalvia: Margaritiferidae). – PLoS ONE 13 (3). https://doi.org/10.1371/journal. pone.0193637.
- Marwaha, Janhavi et al. 2017. Duration of the parasitic phase determines subsequent performance in juvenile freshwater pearl mussels (Margaritifera margaritifera). – Ecology and Evolution 7 (5): 1375–1383. http://hdl.handle.net/1956/17565.
- Marwaha, Janhavi et al. 2019. Host (Salmo trutta) age influences resistance to infestation by freshwater pearl mussel (Margaritifera margaritifera) glochidia. – Parasitology Research 118 (5): 1519–1532. https://doi.org/10.1007/s00436-019-06300-2.
- San Miguel, Eduardo et al. 2004. Growth models and longevity of freshwater pearl mussels (Margaritifera margaritifera). – Spain. Can. J. Zool. 82: 1370–1379. doi: 10.1139/Z04-113.
- Sousa, Ronaldo et al. 2019. A tale of shells and claws: The signal crayfish as a threat to the pearl mussel Margaritifera margaritifera in Europe. – Science of The Total Environment 665: 329–337. https://doi.org/10.1016/j.scitotenv.2019.02.094.
- Taubert, Jens-Eike 2013. Variable development and excystment of freshwater pearl mussel (Margaritifera margaritifera L.) at constant temperature. – Limnologica 43: 319–322. http://dx.doi.org/10.1016/j.limno.2013.01.002.
- Vilbaste, Gustav 1924. Pärlite püüdmisest Eestis. – Loodus 3 (8): 426–428.
- https://www.iucnredlist.org/species/12799/128686456#population. Vaadatud 12.06.2023.
KATRIN KALDMA (1968) on RMK Põlula kalakasvanduse ebapärlikarbispetsialist (projekt LIFE Revives) ja Eesti maaülikooli molekulaarse ökoloogia lektor.