Tekst: Tiiu Kull Fotod: Arto-Randel Servet

Eesti orhideekaitse klubi (EOKK) on tänavuse aasta orhideeks valinud valge tolmpea (Cephalanthera longifolia). Õitsev tolmpea paistab kaugele ja teda on raske teiste liikidega segi ajada. Taime on põnev uurida, sest talle on omased paljunemise eripärad ja varjatud kooselu seentega. Tolmpeade seisundit on Eestis hinnatud ohualtiks, kuna sobivad kasvukohad kipuvad kaduma.
Tolmpea perekond Cephalanthera on ladinakeelse teadusliku nimetuse saanud peakujulise tolmuka järgi: kreekakeelsetest sõnadest kephale ’pea’ ja anthera ’tolmukas’. Maailmas on teada 19 liiki tolmpäid.
Eestis kasvab peale valge tolmpea punane tolmpea. See roosakaspunaste õitega orhideeline õitseb veidi hiljem, aga kasvukoha eelistused on üsna samasugused kui valgel tolmpeal. Õitsevaid taimi on üsna lihtne eristada õite värvi järgi. Keerulisem on määrata punase tolmpea valgeõielist vormi, mida on ka Eestist väga harva leitud. Siis peab vaatama huule kuju, mis punasel tolmpeal on terava, aga valgel tolmpeal tömbi otsaga. Viljuvatel taimedel saab vahet teha ka kupra karvasuse järgi: punasel tolmpeal on kuprad karvased, ent valgel paljad.
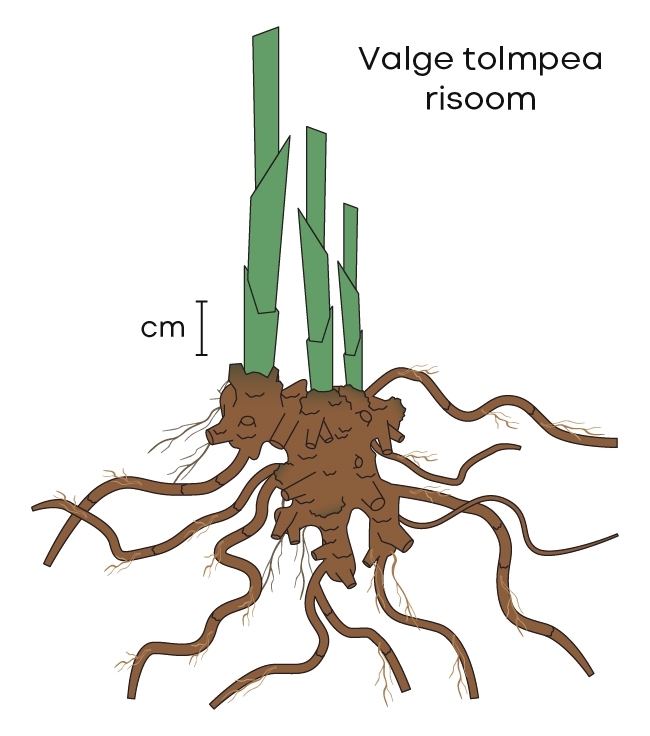
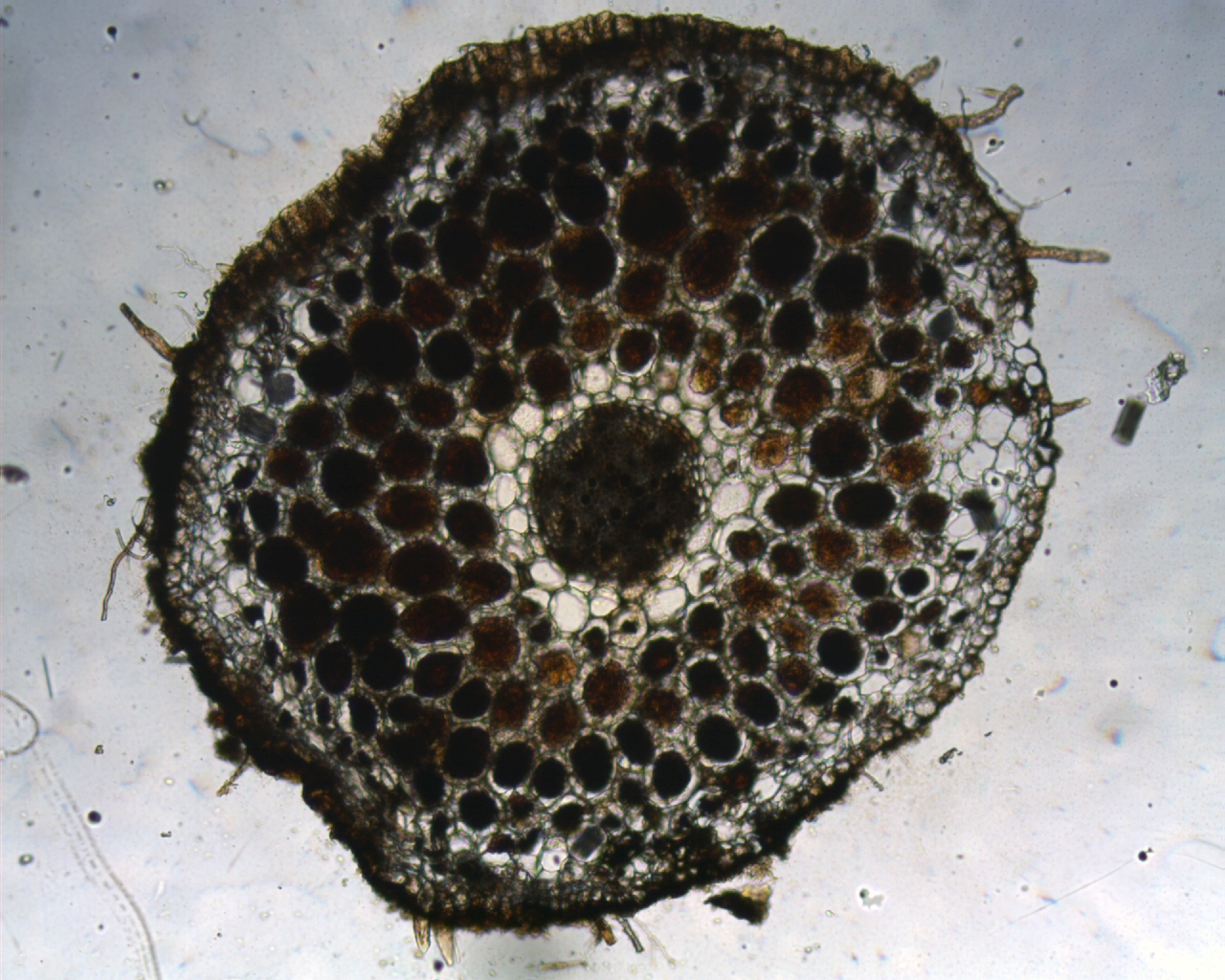
Valge tolmpea vars on püstine, sirge või veidi võnklik ning ümmargune ja kergelt vaoline, lehistunud, alusel asetseb kaks kuni neli labata lehetuppe. Kogu taime kõrgus on tavaliselt 20–50 cm, harva 15–70 cm. Valge tolmpea lehed on pikad ja tugevad, vihmaveerennitaoliselt nõgusad. Neil on ka tugev terav tipp. Risoom on lühike ja püstine (joonis 4). Risoomiharude pungakestest võivad vahel ka võsud areneda. Nii näeme mõnikord looduses mitut valget tolmpeavart tihedas puhmikus koos kasvamas.
tähistatud 2021. aastal Tallinnast Harkust leitud isend
Levik Eestis ja maailmas
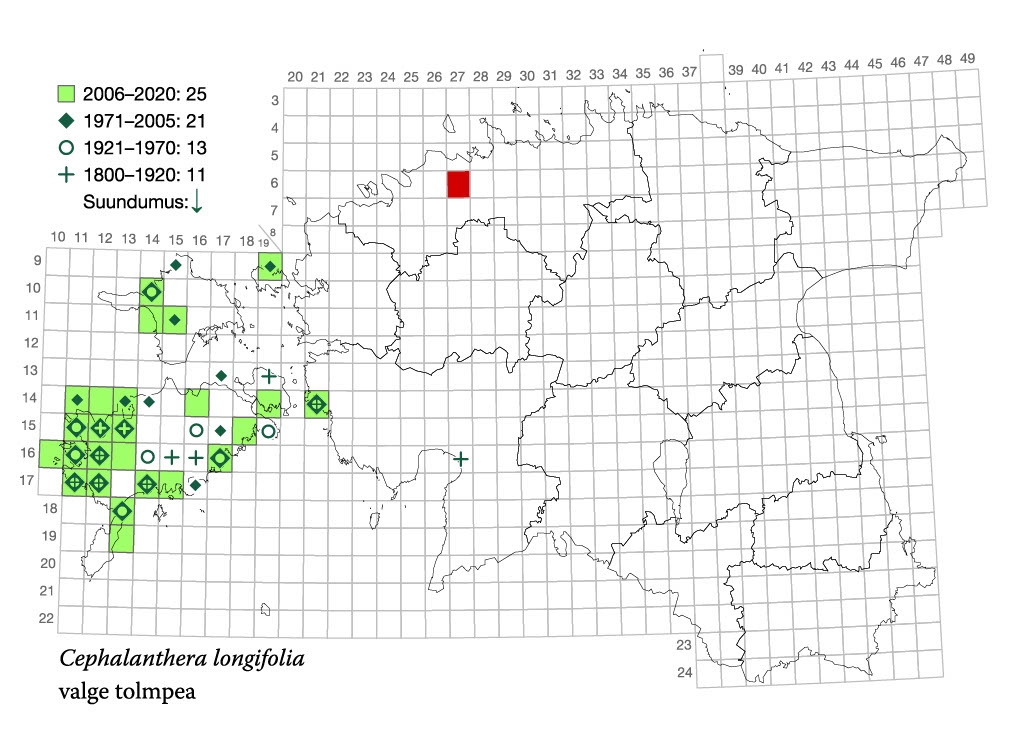
Valge tolmpea põhilised leiukohad paiknevad Lääne-Eesti saartel (joonis 1). Mandri-Eestis on võimalik teda kohata ainult Virtsu ümbruses ja samas lähedal Nehatus. Need leiukohad on botaanikutele teada ammustest aegadest.
2021. aastal leiti üksik valge tolmpea Tallinnast Harku terviseraja äärest, kus ka lähiümbruses sobilikke kasvukohti ei leidu. Taim on seal õitsenud kahel aastal järjest. Tavaliselt pudeneb enamik käpaliste seemneid emataime lähiümbrusesse, kuid nende tolmpeened seemned on võimelised tuulega levima pikkade vahemaade taha.
Loodus pakub alati avastusrõõmu, pealegi ei jõua taimetundjad iial kõiki põõsataguseid ja niidunurki läbi vaadata. Nii võime oletada, et Virtsu ja Tallinna vahel kasvab seni avastamata valgeid tolmpäid veelgi. Seega pakub tänavuse aasta orhidee kindlasti põnevust ka orhideeotsijatele.
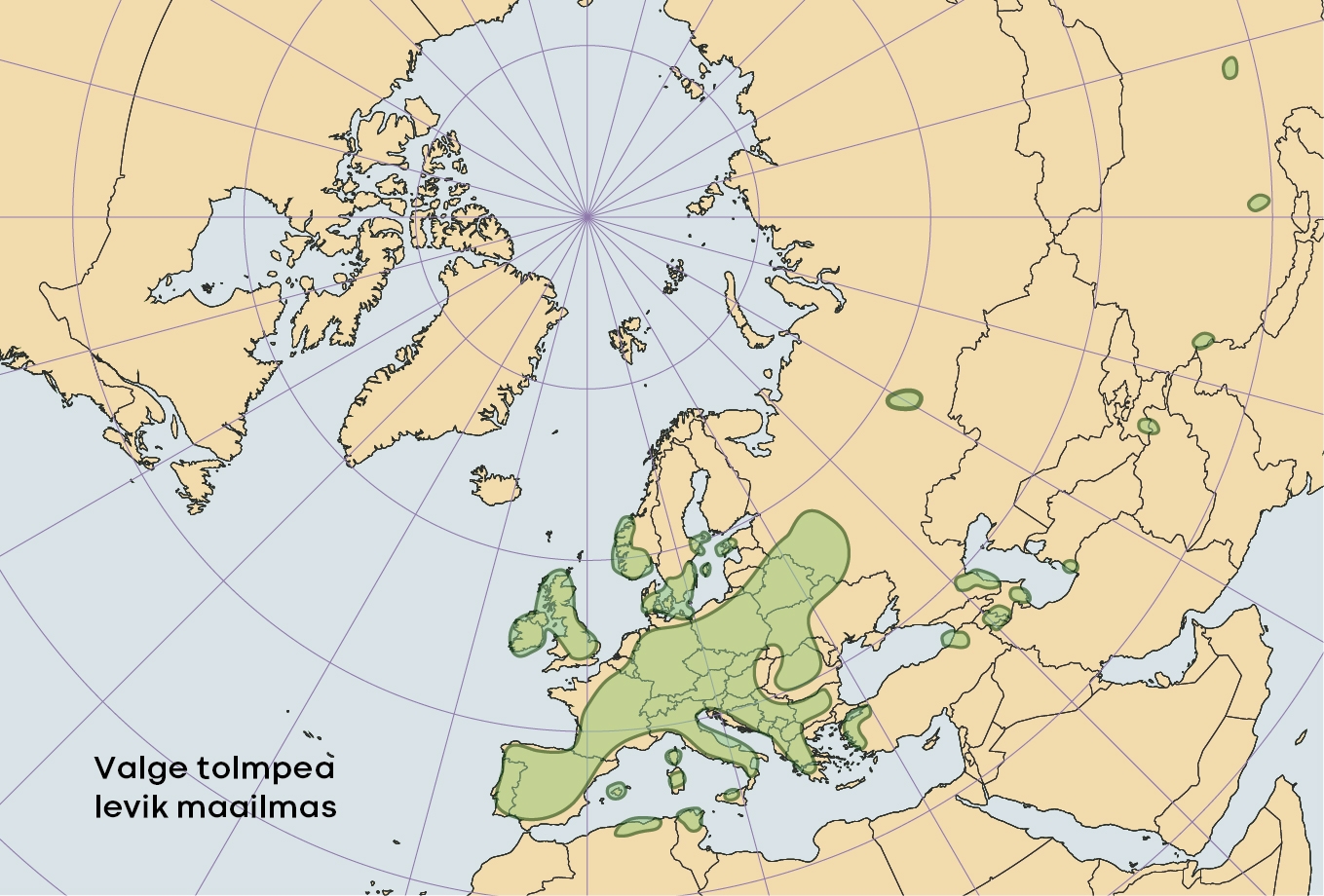
Valge tolmpea on levinud peamiselt Euroopas (joonis 2), lõunas ulatub levila Põhja-Aafrikasse ning idas Iraani ja Himaalajani. Põhja pool läbib areaali piir Skandinaaviat ja Eestit.
Suured õied petavad tolmeldajaid
Valge tolmpea on meie käpalistest üks suuremate õitega: need võivad olla kuni kahe sentimeetri pikkused. Õied on lumivalged, huul aga kollakas. Huul on teistest õiekattelehtedest lühem, umbes ühe sentimeetri pikkune, ja koosneb kahest osast, mis on omavahel ühendatud hingega (joonis 3).
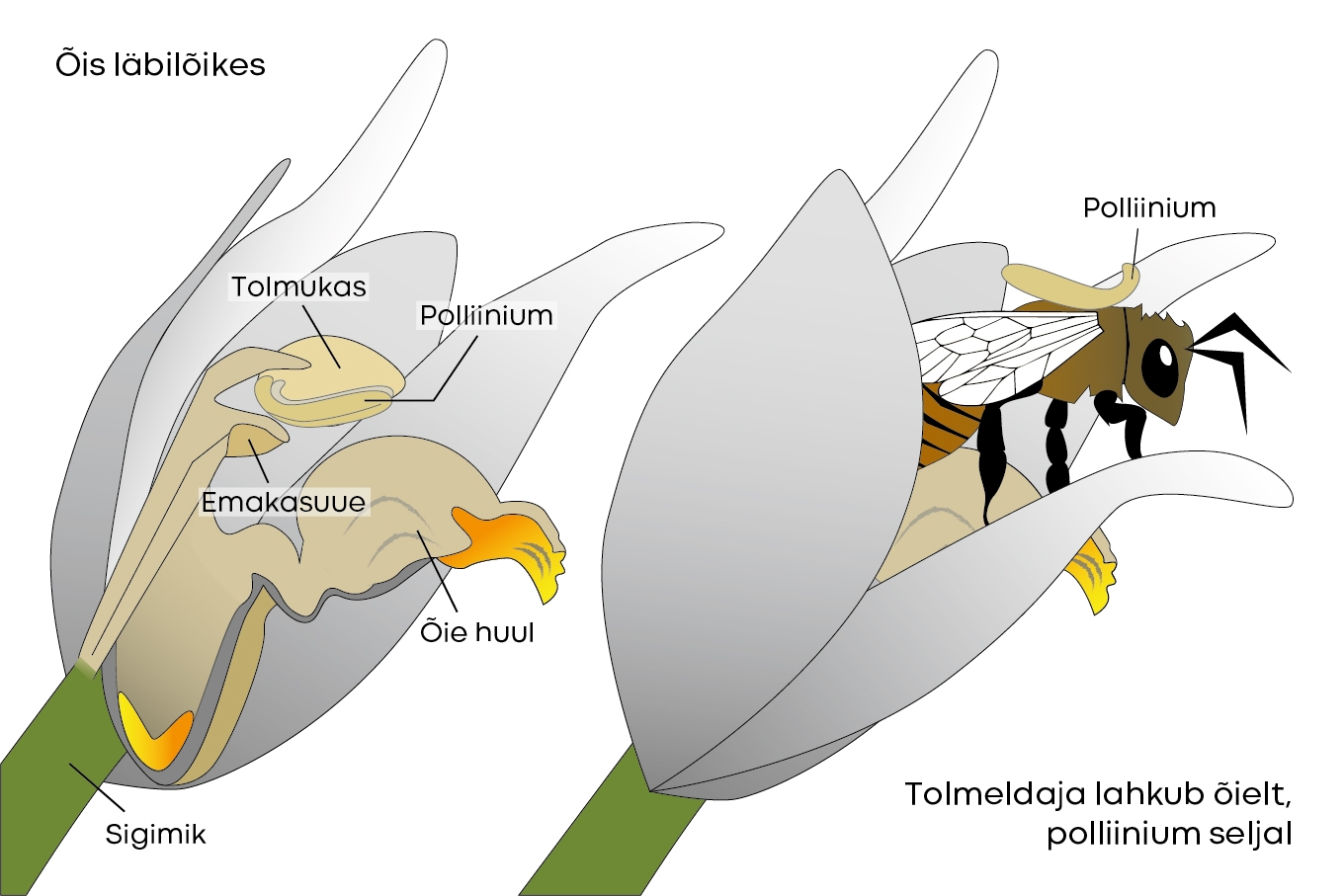
Peakujulises tolmukas asub kaks banaanikujulist polliiniumi, mis kleepuvad õies liikuva putuka seljale ja nii kantakse järgmisele õiele. Tolmuka all paikneb avar emakasuue. Nektarit õies ei leidu, kuid kollane huul köidab ilmselt niigi mesilaste tähelepanu. Peale selle leidub õies lõhnaaineid (pentadekaan ja heptadekaan koos cis-beeta-farneseeniga, mis aitavad putukaid ligi meelitada [2]. Liigi levila lõunaosas on märgatud, et tolmeldajatele väga ligitõmbava salveilehise kiviroosiku (Cistus salviifolius) lähedal viljub valge tolmpea edukamalt [4].
Valget tolmpead tolmeldavad mesilased perekondadest ahasmesilane (Halictus) ja Lasioglossum [3, 4]. Samad mesilased on ka meie teiste käpaliste, näiteks kuldkinga tolmeldajate seas.
Valge tolmpea hakkab õitsema mai lõpus; üks taim õitseb umbes kolm nädalat, üksik õis aga umbes nädala. Jahedamatel aastatel võib mõnda õit näha veel juuli alguses. Üpris hõredas õisikus võib olla kuni paarkümmend õit.
Nagu teistel nektarita petturliikidel, kes tõmbavad tolmeldajaid ligi muude märkidega, on viljumisedukus väike, alla 10%. Viljakuprad on püstised (foto) ja neis areneb umbes 6000 käpalise kohta suhteliselt suurt seemet (1,5 x 0,2 mm) [7].
Nii nagu enamik meie käpalisi, suudab ka valge tolmpea kasvuperioode vahele jätta ja maa all soikeseisundis ebasoodsaid aegu üle elada. Soikeseisund võib kesta kuni kolm aastat. Seetõttu võime samas leiukohas õitsevaid taimi näha aastati eri arvul.
Üleni valged taimed püsivad seentel
Eestis kasvab mitu täiesti seentoidulist orhideed, kellel polegi klorofülli ega lehti, nagu pruunikas pesajuur, lehitu pisikäpp ja kõdu-koralljuur. Vahel harva võib klorofüll puududa ka tavaliselt roheliste lehtedega liikidel.

Saaremaalt on leitud valge tolmpea klorofüllita taimi, kes on üleni valkjad (albiinod) (foto) ega suuda fotosünteesida [9]. Võimalikud on ka vahepealsed variandid, kus lehed on hele-kollakasrohelised. Elus püsivad sellised isendid tänu seentele, kes varustavad neid toitainetega.
Tolmpea perekonnas tuleb ette ka püsivalt klorofüllita liike, näiteks Ameerika ainuke selle perekonna esindaja Cephalanthera austiniae. Itaalias on valge tolmpea seensümbiondid paljud seened, nagu harilik suitsik (Bjerkandera adusta), perekond sametkoorik (Tomentella), Phlebia acerina, sugukonna Sebacinaceae liigid, perekond Tetracladium jt (joonis 5). Seega pole valge tolmpea oma mükoriisse sideme poolest kuigi valiv [8].
Eestist võetud proovid on näidanud valge tolmpea seost tiksikulaadsetega (Helotiales) ja lehternahkiselistega (Thelephoraceae) [1]. Lehternahkiseliste hulka kuulub ka Itaalia uuringutes mainitud sametkoorik, kes on oluline puudega ektomükoriisa moodustaja ja võimaldab lämmastiku ja süsiniku ülekannet puult orhideele. Ühtlasi on see põhjus, miks valge tolmpea kasvab hõredates metsades, kadastikel ja puisniitudel. Läheduses peab olema puid, kuid liiga pimedas kasvukohas õitsevad taimed vähem, mida kinnitavad ka varjutuskatsed [10].
Ülle Püttsepp seiras pikka aega Lääne-Saaremaal kasvanud valge tolmpea populatsiooni, kus leidus ka albiinosid. Ta tõdes, et need olid rohelistega võrreldes väiksemad, neil oli vähem lehti ja õisi ning nende suremus oli suurem, kuid geneetiline erisus puudus [12]. Veidi edukamad olid albiinod siis, kui nad olid soikeseisundi järel maapinnale ilmunud. Arvatavasti toimib neil partnerlus seenega paremini kui roheliste lehtedega isenditel [11]. Seda nähtust võiks vaadelda evolutsioonilise vaheastmena autotroofsuselt mükoheterotroofiale.
Albinootlised taimed on pigmendita aastast aastasse: nad ei ole ühel aastal valged ja teisel rohelised. Taimede klorofüllisisaldus oli vähenenud 99,4%. Mükoriisaseente hulk juurtes suurenes albiinodel umbes kaks korda, kuid seenpartnerid olid samad mis roheliste lehtedega taimedel. Stabiilsete isotoopide koostise analüüs süsiniku kohta näitas, et albiinod sõltusid täielikult seentest, kuid rohelised isendid said seente abil üksnes 33% süsinikust.
Valge tolmpea ja inimtegevus
Valge tolmpea võeti Eestis looduskaitse alla esimeste taimeliikide seas 1936. aastal esimese looduskaitseseadusega. Praegu on ta teise kaitsekategooria liik ja Eesti liikide punase nimestiku ohustatuse hinnangu järgi kategoorias ohualdis. Võrreldes kümne aasta taguse ajaga on valge tolmpea ohustatus suurenenud.
Kõige põhilisem valge tolmpea ohutegur on elupaikade võsastumine. Loometsad ja puisniidud on meil kujunenud valgusküllaseks suuresti karjatamise ja mõõduka majandamise oludes. Kui selline maatarvitus on lõppenud, kuivab muld ajuti läbi ning looduslikud muutused ja taasmetsastumine kestab sageli väga pikka aega. Vanemates loometsades võivad kasvukohad püsida kaua aega suhteliselt avatuna ja valgusküllasena ka inimese pideva sekkumiseta. Nii inimtekkelistel kui ka looduslikel häiringutel võib olla küllaltki suur tähtsus. Mõõduka kasutuse korral püsivad loometsades avatuna ka liikumisteed ja metsasihid, moodustades tolmpeale sidusa kasvukohtade võrgustiku.

Ohtlik on ka kinnisvaraarendus, sest loopealsed ja kadastikud on selle ihaldatud alad. Kui taimed on kadunud kasvukoha kinnikasvamise tagajärjel, siis säilib vähemalt mõnda aega lootus, et kui elupaik taastada, taastub alal säilinud seemnepanga kaudu ka populatsioon. Seevastu arendus hävitab kasvukoha enamasti jäädavalt ning ühtlasi killustab ehitustöö ka tolmeldajate levikut.
Metssead võivad tuhnides kahjustada juuremugulatega käpaliste kõrval ka valge tolmpea populatsioone, mistõttu peaks sigade arvukust kontrolli all hoidma; kindlasti ei tohi orhideerikastel aladel korraldada lisasöötmist.
Tiiu Kull (1958) on botaanik ja orhideeuurija, Eesti maaülikooli emeriitprofessor, Eesti orhideekaitse klubi asutajaliige.
1. Abadie, Jean-Claude et al. 2006. Cephalanthera longifolia (Neottieae, Orchidaceae) is mixotrophic: a comparative study between green and nonphotosynthetic individuals. – Botany 84 (9): 1462–1477.
2. D’Auria, Maurizio et al. 2021. Aroma components of Cephalanthera orchids. – Natural product research 35 (1): 174–177.
3. Claessens, Jean; Kleynen, Jacques 2013. The pollination of European orchids: part 2: Cypripedium and Cephalanthera. – Journal of the Hardy Orchid Society 10: 114–120.
4. Dafni, Amots; Yariv, Ivri 1981. The flower biology of Cephalanthera longifolia (Orchidaceae) – pollen imitation and facultative floral mimicry. – Plant Systematics and Evolution 137: 229–240.
5. Hultén, Eric; Fries, Magnus 1986. Atlas of North European Vascular Plants North of the Tropic of Cancer. Koeltz Scientific Books, Königstein.
6. Kukk, Toomas jt 2020. Eesti taimede levikuatlas 2020. Pärandkoosluste kaite ühing, Eesti maaülikooli põllumajandus- ja keskkonnainstituut, Tartu.
7. Mrkvicka, Alexander C. 1994. Anatomie und Morphologie der Samen heimischer Orchideenarten. – Journal Europäischer Orchideen 26 (2): 168–314.
8. Pecoraro, Lorenzo et al. 2017. Fungal diversity and specificity in Cephalanthera damasonium and C. longifolia (Orchidaceae) mycorrhizas. – Journal of Systematics and Evolution 55 (2): 158–169.
9. Püttsepp, Ülle 1993. Kahvatu orhidee. – Eesti Loodus 44 (11/12): 357.
10. Shefferson, Richard P. et al. 2006. Demographic response to shading and defoliation in two woodland orchids. – Folia Geobotanica 41: 95–106.
11. Shefferson, Richard P. et al. 2016. Demographic shifts related to mycoheterotrophy and their fitness impacts in two Cephalanthera species. – Ecology 97 (6): 1452–1462.
12. Tranchida‐Lombardo, Valentina et al. 2010. Spatial repartition and genetic relationship of green and albino individuals in mixed populations of Cephalanthera orchids. – Plant Biology 12 (4): 659–667.